Biology Textbook Fraud
Use this fraud test on your own textbooks
How homology is argued and defined
Informed scientists speak about homology
|
Textbook Fraud |
|
|
|
|
|
|
|
|
|
|
|
|
Biology Textbook Fraud Use this fraud test on your own textbooks How homology is argued and defined Informed scientists speak about homology |
|
|
|
|
|
|
|
|
|
|
How Evolutionists Argue Homology |
|
|
|
|
|
Darwin |
"We have seen that the members of the same class, independently of their habits of life, resemble each other in the general plan of their organization. This resemblance is often expressed by the term "unity of type" - or by saying that the several parts and organs in the different species of the class are homologous. The whole subject is included under the general term of Morphology. This is one of the most interesting departments of natural history, and may almost be said to be its very soul. What can be more curious than that the hand of a man, formed for grasping, that of a mole for digging, the leg of the horse, the paddle of the porpoise, and the wing of the bat should all be constructed on the same pattern, and should include similar bones, in the same relative positions? How curious it is, to give a subordinate though striking instance, that the hind-feet of the kangaroo, which are so well fitted for bounding over the open plains, - those of the climbing, leaf eating koala, equally well fitted for grasping the branches of trees, -those of the ground -dwelling, insect or root-eating, bandicoots, - and those of some other Australian marsupials, - should all be constructed on the same extraordinary type, namely with the bones of the second and third digits extremely slender and enveloped within the same skin, so that they appear like a single toe furnished with two claws. Notwithstanding this similarity of pattern, it is obvious that the hind feet of these several animals are used for as widely different purposes as it is possible to conceive. The case is rendered all the more striking by the American oppossums, which follow nearly the same habits of life as some of their Australian relatives, having feet constructed on the ordinary plan. Professor Flower, from whom these statements are taken, remarks in conclusion: "We may call this conformity of type, without getting much nearer to an explanation of the phenomenon"; and he then adds "but is it not powerfully suggestive of true relation-ship, of inheritance from a common ancestor?" (The Origin of Species, Charles Darwin, 1872, 6th ed, 1962, Collier Books, New York p 434-5) |
|
|
|
|
Encyclopaedia Britannica |
"It must be stressed that Darwin himself never claimed to provide proof of evolution or of the origin of species, what he did claim was that if evolution has occurred, a number of otherwise inexplicable facts are readily explained. The evidence for evolution was therefore indirect.... The indirect evident for evolution is based primarily on the significance of similarities found in different organisms. . . . The similarity of plan is easily explicable if all descended with modification from a common ancestor, by evolution, and the term homologous is used to denote corresponding structures formed in this way. . . . In vertebrate animals, the skeleton of the forelimb is a splendid example of homology, in the bones of the upper arm, forearm, wrist, hand, and fingers, all of which can be matched, bone for bone, in rat, dog, horse, bat, mole, porpoise, or man. The example is all the more telling because the bones have become modified in adaptation to different modes of life but have retained the same fundamental plan of structure, inherited from a common ancestor." (Encyclopaedia Britannica (1981) 15th ed, Encyclopaedia Britannica. Inc, Chicago, Macropaedia vol 7, p 8) |
|
|
|
Similarity in design (homology) is logically attributed to a single creator/designer unless it can be shown otherwise. For any evolutionist to reject a designer because of similarities, is contrary to the most basic forms of human reasoning. If a person comes across a single rock laying at the bottom of a cliff, they would logically conclude that it rests there by chance processes. Even if it had been laid there as some kind of "modern artwork" of a designer, the first conclusion would not include a designer. But if the same person came across a stone house at the bottom of the same cliff, they would logically conclude a designer was involved. If would be illogical for anyone to conclude the stone house was the result of random chance processes. There is not a man alive who finds a watch on the ground who would conclude the watch "just happened without a designer". But believe it or not, evolutionists are just that illogical! Richard Dawkins wrote a book called "The Blind Watchmaker", where he insults the intelligence of all humans by suggesting that, yes, watches can result from random chance processes under the direction of his god called "natural selection". We find it sad that such a brilliant man can inflict upon himself the ridicule of the planet rather than admit the obvious, that life on earth is the result of a designer. (God)
General Motors makes several different families of automobiles. You have small compact, midsize and full size luxury. Within each of these families, there are variations of specific models. For example, Al Haas, an auto reviewer wrote: "The Regal is part of a family of new General Motors midsize sedans that includes the Oldsmobile Intrigue and the Pontiac Grand Prix. Like the Buick, Olds and Pontiac midsize sedans that debuted a decade ago, these cars are very similar structurally and mechanically. What's interesting is that, unlike the three GM midsizers introduced 10 years ago, these cars are dissimilar in appearance and character. So, instead of being roundly booed by the media and industry analysts for their sameness, as their ancestors were, they are being hailed for the extent to which they have been differentiated in appearance, personality and driving manners - and in the kinds of customers they appeal to." (Philadelphia Inquirer, Al Haas, Automotive Reviewer, April 96.) Notice the use of the words, "family" and "ancestors" by this auto reviewer. Similarity in design (homology) is automatically attributed to a single creator/designer unless it can be shown otherwise. But the evolutionist quips back, "How do you explain similarity in auto design with 10 different auto manufacturers? That is similarity without a common designer like evolution." But, homology of design with different auto designers (manufacturers) makes the case for creation even stronger. You see the designers realized that there were "best ways" to do certain things in our physical world. Having tried and tested all alternatives, they all settled on the same basic design because it was simply the best. That explains why God chose to use common design features for many forms of life. Similarity ('homology') is not an absolute indication of common ancestry (Evolution) but certainly points to a common designer (creation).
|
de Beer Identifies Some Problems Sir Gavin says that there is no doubt whatever that the forelimb in the newt and the lizard and the arm of man are strictly homologous, inherited with modification from the pectoral fin of fishes 500 million years ago. The elbow and wrist joints are identical, and their hands end in five fingers. The bones and muscles also correspond. The shock comes when a careful study of their comparative anatomy reveals that they do not occupy the same positions in the body. "The limbs of vertebrates are always formed from material that is contributed from several adjacent segments of the trunk. In the newt, the forelimb is formed from trunk segments 2, 3, 4, and 5; in the lizard from 6, 7, 8, and 9; in man from trunk segments 13 to 18 inclusive." He explains how this is determined embryologically and anatomically. Embryology was de Beer's basis for questioning the validity of homology as evidence for evolution. He said that progress in early embryology made such strides there are two levels on which the relations of homology and embryology can be studied:
At level one, de Beer says that the prospective fates of embryonic structures are well known, and they can be traced back to their place on the egg or young embryo of related species. [de Beer, p. 13] They can be observed and proved by experiment. In other words, the wrongful application of homology by evolutionists has been debunked by the scientific method. Sir Gavin gives us some examples of this logical approach: use of the scientific method to evaluate evolutionary claims. The alimentary canal in vertebrates can form from:
It is evident that homologous structures are known to develop from varying locations in the egg or embryo, depending on the species. Therefore, says de Beer, if the origin of the homologous structures is not from the same part of the egg or position of the cells in the embryo, then they are not genetically related because they are not under the control of homologous genes. If they are not genetically related, they are not the result of descent with modification from a common ancestor. He draws the same conclusion from a different set of observations. In one species of Polygordius, a primitive worm (primitive is subjective), the trunk of the future worm develops inside the body of the larva, while in another species, the trunk develops outside the larva as a worm-like extension. The adults of both species are virtually indistinguishable. Similar differences are seen in the development of certain mollusks, although the adults are nearly identical. At level two de Beer studied homologous relations by inducing embryonic tissues to differentiate by diffusing substances from a master structure called an organizer. One study showed that the dorsal lip of a newt embryo can be grafted anywhere into the body of another embryo and will induce the surrounding tissues to differentiate into all the structures characteristic of a vertebrate embryo. If these tissues had been left undisturbed, they would have differentiated into entirely different structures (notochord, segmental muscle plates, kidney tubules spinal cord, brain with eyes, etc.), proving that the nature of a structure does not depend on the place of origin. This should be very disturbing to those who wish to apply homology to the battery of evidences for evolution. Homologous structures should originate at the same places in all descendants of a common ancestor, otherwise the homology has nothing to do with descent. In another example, de Beer says that if the optic cup is removed in one species of frog the eye lens will not develop, while in another closely related species the lens does develop. He says it cannot be doubted that the lenses of these two species are homologous, yet they differ completely in their mechanism of determination and differentiation. The genes involved are not homologous. In summary, he says: "It is now clear that the pride with which it was assumed that the inheritance of homologous structures from a common ancestor explained homology was misplaced; for such inheritance cannot be ascribed to identity of genes. The attempt to find 'homologous' genes, except in closely related species, has been given up as hopeless." He understands the logical absurdities, quoting S.C. Harland who said that the genes coding for the homologous structures must have become wholly altered during the evolutionary process! This should alarm geneticists, computer scientists, and linguists. It simply doesn't make sense, and de Beer says so: "But if it is true that through the genetic code, genes code for enzymes that synthesize proteins which are responsible (in a manner still unknown in embryology) for the differentiation of the various parts of their normal manner, what mechanism can it be that results in the production of homologous organs, the same 'patterns' in spite of their not being controlled by the same genes? I asked this question in 1938, and it has not been answered." |
Similar structures from different ancestry (eye of mouse and octopus)
Evolutionary biologist Ernst Mayr defined homology as, "A feature in two or more taxa is homologous when it is derived from the same (or a corresponding) feature of their common ancestor." (Mayr, 1982)
As the title of his book indicates, Berra's primary purpose is to show that living organisms are the result of naturalistic evolution rather than supernatural design. Structural similarities among automobiles, however, - even similarities between older and newer models (which Berra calls "descent with modification"), - are due to construction according to pre-existing patterns, i.e., to design. Ironically, therefore, Berra's analogy shows that phylogenetic homology is not sufficient to exclude design-based explanations. In order to demonstrate naturalistic evolution, it is necessary to show that the mechanism by which organisms are constructed (unlike the mechanism by which automobiles are constructed) does not involve design.
One could simply postulate that the mechanism of biological evolution is naturalistic, arguing that the postulate is justified because science is limited to studying natural mechanisms. Although such a philosophical move may seem very reasonable, however, it compromises the status of evolutionary biology as an objective science. Asserting that something is objectively true implies that it is based on empirical evidence, not merely assumed a priori on philosophical grounds. A methodological exclusion of design-based explanations constitutes a limitation on one's discipline, not a description of objective reality. If evolutionary biologists want to show that the actual mechanism of evolution does not involve supernatural design, they cannot merely exclude the possibility a priori, but must take the more difficult approach of proposing and corroborating a naturalistic alternative.
This alternative must account naturalistically for what evolutionary biologist Leigh Van Valen has called "continuity of information." (Van Valen, 1982) According to Van Valen, homologous features are produced during the development of each individual organism by information which has been inherited, with modification, from the organism's ancestors. Thus the first step toward understanding the mechanism of evolution would be to determine the nature of the information which controls the development of the embryo.
One possibility is that this information is encoded in the organism's genes. In the 1930's, the synthesis of Darwin's theory and population genetics explained evolution as a change in gene frequencies, and several decades later the discovery of the structure and function of DNA extended this explanation to the molecular level.
According to the neo-Darwinian synthesis, a genetic program encoded in DNA directs embryonic development; the process of reproduction transmits this program to subsequent generations, but mutations in the DNA sometimes modify it ("descent with modification"); thus descendants of the original organism may possess structures which are similar but not identical ("homologies"). No design is required, so the explanation is thoroughly naturalistic. By 1970, molecular biologist Jacques Monod felt justified in announcing that "the mechanism of Darwinism is at last securely founded," and that as a consequence "man has to understand that he is a mere accident." (quoted in Judson, 1980, p. 217)
Efforts to correlate evolution with changes in gene frequencies, however, have not been very successful. Detailed studies at the molecular level fail to demonstrate the expected correspondence between changes in gene products and the sorts of organismal changes which constitute the "stuff of evolution." (Lewontin, 1974, p. 160). According to Rudolf Raff and Thomas Kaufman, evolution by DNA mutations "is largely uncoupled from morphological evolution;" the "most spectacular" example of this is the morphological dissimilarity of humans and chimpanzees despite a 99% similarity in their DNA. (Raff and Kaufman, 1983, pp. 67, 78).
Some biologists have proposed that the remaining 1% consists of "regulatory genes" which have such profound effects on development that a few mutations in them could account for dramatic differences. For example, mutations in homeotic genes can transform a fly's antenna into a leg, or produce two pairs of wings where there would normally be only one, or cause eyes to develop on a fly's leg. Furthermore, genes similar to the homeotic genes of flies have been found in most other types of animals, including mammals. Based on the profound developmental effects and almost universal occurrence of such genes, biologist Eric Davidson and his colleagues recently wrote that "novel morphological forms in animal evolution result from changes in genetically encoded programs of developmental regulation." (Davidson et al., 1995, p. 1319)
According to this view, homologous features are programmed by similar genes. Assuming that genes with similar sequences are unlikely to originate independently through random mutations, sequence similarity would indicate common ancestry. Features produced by similar sequences could then be inferred to be phylogenetically homologous.
The very universality of homeotic genes, however, raises a serious problem for this view. Although mice have a gene very similar to the one that can transform a fly's antenna into a leg (Antennapedia), mice do not have antennae, and their corresponding gene affects the hindbrain; and although mice and flies share a similar gene which affects eye development (eyeless), the fly's multifaceted eye is profoundly different from a mouse's camera-like eye. In both cases (Antennapedia and eyeless), similar homeotic genes affect the development of structures which are non-homologous by either the classical morphological definition or the post-Darwinian phylogenetic definition. If similar genes can "determine" such radically different structures, then those genes aren't really determining structure at all. Instead, they appear to be functioning as binary switches between alternate developmental fates, with the information for the resulting structures residing elsewhere. (Wells, 1996)
Not only are non-homologous structures produced by organisms with supposedly homologous genes, but organisms with different genes can also produce similar structures. The most famous examples involves the genes, mentioned above, which affect wing and eye development in flies. Fly embryos with a normal gene for wing development, when treated with ether, can be induced to grow a second pair, just as though they possessed the mutant form of the gene. (For a review, see Hall, 1992) Flies with a mutant form of the eye gene fail to develop eyes; but if eyeless flies are bred for many generations, some of their descendants will develop eyes even though they still possess the mutant form of the gene. Such anomalies led embryologist Gavin de Beer to conclude that "homologous structures need not be controlled by identical genes," and that "the inheritance of homologous structures from a common ancestor...cannot be ascribed to identity of genes." (de Beer, 1971, pp. 15-16)
The underlying assumption that a genetic program directs embryonic development has been seriously questioned by developmental biologists. (For a review, see Wells, 1992) Sydney Brenner, who originally proposed genetic programs in 1970, repudiated the idea when he realized that the information required to specify the neural connections of even a simple worm far exceeds the information content of its DNA. (Brenner, 1973) A decade later, developmental biologist Brian Goodwin noted that "genes are responsible for determining which molecules an organism can produce," but "the molecular composition of organisms does not, in general, determine their form." (Goodwin, 1985, p. 32) And in a 1990 critique of the notion of genetic programs, H.F. Nijhout concluded that "the only strictly correct view of the function of genes is that they supply cells, and ultimately organisms, with chemical materials." (Nijhout, 1990, p. 444)
Clearly, the genetic explanation for homology is inadequate. As an alternative, some biologists have suggested that homology results from complex developmental mechanisms which are not reducible to a genetic program.
Since homologies cannot be explained by equating developmental information with DNA sequences, some biologists have attempted to explain it by attributing it to similar developmental pathways. Although DNA determines the amino acid sequence of proteins essential for development, such pathways also involve other factors, such as the localization of cytoplasmic constituents in the egg cell, physical constraints resulting from the size of the embryo, and so on. (Wells, 1992)
Efforts to correlate homology with developmental pathways, however, have been uniformly unsuccessful. First, similar developmental pathways may produce very dissimilar features. At the molecular level, it is well known that virtually identical inducers may participate in the development of non-homologous structures in different animals. (Gilbert, 1994) At the multicellular level, the pattern of embryonic cell movements which generates body form in birds also generates body form in a few species of frogs. (Elinson, 1987) And even at the organismal level, morphologically indistinguishable larvae may develop into completely different species. (de Beer, 1958) Clearly, similar developmental pathways may produce dissimilar results.
Second, and more dramatically, similar features are often produced by very different developmental pathways. No one doubts that the gut is homologous throughout the vertebrates, yet the gut forms from different embryonic cells in different vertebrates. The neural tube, embryonic precursor of the spinal cord, is regarded as homologous throughout the chordates, yet in some its formation depends on induction by the underlying notochord while in others it does not. (Gilbert, 1994) Evidently, "structures can owe their origin to different methods of induction without forfeiting their homology." (de Beer, 1958, p. 151) Indeed, as developmental biologist Pere Alberch noted in 1985, it is "the rule rather than the exception" that "homologous structures form from distinctly dissimilar initial states." (Alberch, 1985, p. 51)
Production of similar forms from dissimilar pathways is also common at later stages of development. Many types of animals pass through a larval stage on their way to adulthood, a phenomenon known as indirect development. For example, most frogs begin life as swimming tadpoles, and only later metamorphose into four-legged animals. There are many species of frogs, however, which bypass the larval stage and develop directly. Remarkably, the adults of some of these direct developers are almost indistinguishable from the adults of sister species which develop indirectly. In other words, very similar frogs can be produced by direct and indirect development, even though the pathways are obviously radically different. The same phenomenon is common among sea urchins and ascidians. (Raff, 1996)
Even the classic example of vertebrate limbs shows that homology cannot be explained by similarities in developmental pathways. Skeletal patterns in vertebrate limbs are initially laid down in the form of cartilage condensations, which later ossify into bone. The sequence of cartilage condensation is the developmental pathway which determines the future pattern of bones in the limb. Yet similar bone patterns in different species (i.e., homologies) arise from different sequences of cartilage condensation. (Shubin, 1991) In the words of biologist Richard Hinchliffe: "Embryology does not contribute to comparative morphology by providing evidence of limb homology in the form of an unchanging pattern of condensation common to all tetrapod limbs." (Hinchliffe, 1990, p. 121)
So embryology has not solved the problem of homology. In 1958, Gavin de Beer observed that "correspondence between homologous structures cannot be pressed back to similarity of position of the cells in the embryo, or of the parts of the egg out of which the structures are ultimately composed, or of developmental mechanisms by which they are formed." (de Beer, 1958, p. 152) Subsequent research has overwhelmingly confirmed the correctness of de Beer's observation. Homology, whether defined morphologically or phylogenetically, cannot be attributed to similar developmental pathways any more than it can be attributed to similar genes. So far, the naturalistic mechanisms proposed to explain homology do not fit the evidence.
In 1802, William Paley wrote that someone crossing a heath and finding a stone could reasonably attribute its presence to purposeless natural causes. Upon finding a watch, however, and seeing that "its several parts are framed and put together for a purpose," one could conclude that the watch had been designed. By analogy, Paley argued, one could also conclude that living things are designed. (Paley, 1802, p. 2) In 1859, Charles Darwin argued that living things are more like Paley's stone than Paley's watch, and claimed that everything which Paley attributed to design could be accounted for naturalistically, by descent with modification.
As Berra's automobile analogy shows, however, descent with modification is not enough to exclude design. It is necessary, in addition, to show that the mechanism of descent with modification is thoroughly naturalistic. Darwin thought he had done this with his theory of natural selection, but as the problem of homology demonstrates, he failed to accomplish his goal.
Diverse organisms possess homologous features. Homology may or may not be due to inheritance from a common ancestor, but it is definitely NOT due to similarity of genes or similarity of developmental pathways. In 1971, Gavin de Beer wrote: "What mechanism can it be that results in the production of homologous organs, the same 'patterns', in spite of their not being controlled by the same genes? I asked this question in 1938, and it has not been answered." (de Beer, 1971, p.16) Twenty-five years later, the question still has not been answered.
Without a naturalistic mechanism to account for homology, however, Darwinian evolution cannot claim to have demonstrated scientifically that living things are undesigned, and the possibility remains that homologies are patterned after idealized archetypes. Without a demonstrated mechanism, naturalistic biologists are left with only one alternative: exclude design a priori, on philosophical grounds. This exclusion could be taken as a statement that supernatural design does not exist, or it could be taken as a statement that supernatural design is beyond the reach of empirical science. The first is a theological statement, and warrants a theological response. The second is a methodological limitation which cannot be logically extrapolated to a limitation on reality. In other words, a scientist who makes the first move is engaging in theological disputation, while a scientist who makes the second is declining to investigate a possible aspect of reality.
Unfortunately, many biologists make both moves, but fail to distinguish logically between them. While justifying their exclusion of supernatural design on methodological grounds, they act as though science has disproved its existence by providing a naturalistic explanation for homology. When confronted with the fact that science has failed in this regard, they reaffirm their methodological commitment and express faith that a naturalistic mechanism will someday be discovered.
And perhaps it will. But what if living things really are designed? Someone who finds a watch on the ground, and wants to investigate its origin, would be mistaken to rule out design a priori. Having already jumped to the wrong conclusion, that person might go on to waste an entire lifetime dabbling in spurious explanations. If science is truth-seeking, then this is a strange way to do science.
According to an old joke, a passer-by walks up to a drunk stumbling around under a street light. The passer-by asks the drunk what he's doing, and the drunk replies, "Looking for my watch." "Oh, did you lose it here?" asks the passer-by. "No," the drunk replies, "I lost it across the street, but there's no light over there!" Letting naturalistic philosophical assumptions limit one's search for the cause of homology may not be the best way to study living things.
Alberch, Pere (1985). "Problems with the Interpretation of Developmental Sequences," Systematic Zoology 34 (1): 46-58
Berra, Tim M. (1990). Evolution and the Myth of Creationism. Stanford, CA: Stanford University Press.
Bowler, Peter J. (1989). Evolution: The History of an Idea. Revised edition. Berkeley: University of California Press.
Brenner, Sydney (1973). "The Genetics of Behaviour," British Medical Bulletin 29: 269-271.
Davidson, E. H., Peterson, K. J. and Cameron, R. A. (1995). "Origin of Bilaterian Body Plans: Evolution of Developmental Regulatory Mechanisms," Science 270: 1319-1325.
de Beer, Gavin (1958). Embryos and Ancestors, 3rd ed. Oxford: Clarendon Press.
de Beer, Gavin (1971). Homology: An Unsolved Problem. London: Oxford University Press.
Elinson, Richard P. (1987). "Change in Developmental Patterns: Embryos of Amphibians with Large Eggs." In Rudolf A. Raff and Elizabeth C. Raff, eds., Development as an Evolutionary Process, vol. 8, pp. 1-21. New York: Alan R. Liss.
Gilbert, Scott F. (1994). Developmental Biology, 4th ed. Sunderland, MA.: Sinauer Associates.
Goodwin, Brian C. (1985). "What Are the Causes of Morphogenesis?" Bioessays 3: 32-36.
Hall, Brian K. (1992). Evolutionary Developmental Biology. London: Chapman & Hall.
Hinchliffe, Richard (1990). "Towards a Homology of Process: Evolutionary Implications of Experimental Studies on the Generation of Skeletal Pattern in Avian Limb Development." In J. Maynard Smith and G. Vida, eds., Organizational Constraints on the Dynamics of Evolution, pp. 119-131. Manchester, UK: Manchester University Press.
Judson, Horace Freeland (1980). The Eighth Day of Creation. New York: Simon & Schuster.
Lewontin, R.C. (1974). The Genetic Basis of Evolutionary Change. New York: Columbia University Press.
Mayr, Ernst (1982). The Growth of Biological Thought. Cambridge, MA: Belknap Press.
Nijhout, H.F. (1990). "Metaphors and the Role of Genes in Development," Bioessays 12: 441-446.
Paley, William (1802). Natural Theology. Reprinted in 1972. Houston, TX: St. Thomas Press,.
Panchen, Alec L. (1994). "Richard Owen and the Concept of Homology." In Brian K. Hall, ed., Homology: The Hierarchical Basis of Comparative Biology. San Diego: Academic Press, pp. 21-62.
Raff, Rudolf A. (1996). The Shape of Life: Genes, Development, and the Evolution of Animal Form. Chicago: The University of Chicago Press.
Raff, Rudolf A. and Kaufman, Thomas C. (1983). Embryos, Genes, and Evolution. New York: Macmillan.
Shubin, Neil H. (1991). "The Implications of 'The Bauplan' for Development and Evolution of the Tetrapod Limb." In J.R. Hinchliffe, J.M. Hurle, and D. Summerbell, eds., Developmental Patterning of the Vertebrate Limb, pp. 411-421. New York: Plenum Press.
Van Valen, Leigh M. (1982). "Homology and Causes." Journal of Morphology 173: 305-312.
Wagner, Guenter (1989). "The Biological Homology Concept," Annual Review of Ecology and Systematics 20: 51-69.
Wells, Jonathan (1992). "The History and Limits of Genetic Engineering," International Journal on the Unity of the Sciences 5: 137-150.
Wells, Jonathan (1996). "Unseating Naturalism: Recent Insights from Developmental Biology." Presented at a conference on Mere Creation: Reclaiming the Book of Nature, Biola University, Los Angeles.

|
Use this fraud test on your own text books: We charge any school textbook with fraud and gross misrepresentation if it:
|
|
|
|
|
|
|
Proof that textbooks in use this school year engage in classroom fraud. |
|
|
|
|
|
|
|
|
|
|
What informed scientists say about Homology |
|
|
|
|
The Fraud Exposed |
|
|
|
|
|
Yolk sac in humans |
"Induction by fibroblast growth factors of mesoderm during gastrulation leads to blood-forming tissue, including angioblasts & hemopoietic cells, that together constitute the blood islands of the yolk sac." (Vasculogenesis, Werner Risau & Ingo Flamme, Annual Review of Cell Division Biology, 1995, p. 73)
As a grad student at the U. of Northern Colorado (I studied under 1 of the top parasitologists in the nation, the late G.D. Schmidt, ('Foundations of Parasitology'), In the mid 80s I gave a presentation to a grad class that included 2 professors. During my presentation, I referred to the blood sac of man ("yolk sac") several times. I was fascinated that I was not challenged when using this title. It's because they knew the blood sac has blood cells in it, not yolk. It's called a "yolk sac" only because, for example turtles & chicks, it does have yolk in it for nutrition for the developing embryo.
In man the y.s. only looks like a y.s., but, of course, it contains no yolk (Figure 5-12 of Hildebrand, 3rd ed)- that's why macroevolutionists call it a yolk sac - go figure. In the meantime, science says the function is different (i.e. it contains small clumps or cords of cells, called blood islands according to embryologist Hart - so much for the person who wanted a "design explanation" for the eutherian "empty" yolk sac). Although macroevolutionists see the yolk sac of most mammals as 'rudimentary vestiges', just see how long the embryo lives wi/o one. The yolk-sac placenta of marsupials ("lower mammals") is needed for gaseous exchange to occur. As to the nine families of snakes & lizards that have placentas, so what? Some mammals lay eggs. When it comes to the "placenta" (you were right to place the word in quotation marks!) in a few species of fish, there is no no true placental attachment. It would be interesting to formulate an experiment that would remove the "placenta" from this developmental process & see what happens. I would just bet the embryos die. I'm disturbed that these professors would throw the word proof around so freely. They should know that it is very difficult to prove anything. When it comes to functional structures we possess "proving" our alleged macroevolutionary history - well, that's a stretch cautious scientists would not be prepared to take. Where do fish & reptiles get their first blood from if the y.s. is used for nutrition? Embryologists find that at various stages of development of the embryo blood comes from a several sources (e.g. embryonic liver). While much is made of this alleged evidence in favor of macroevolution, I'm reminded what evolutionist Dr. A.G. Fisher said in 1998: "Both the origin of life and the origin of the major groups of animals remain unknown." Ironically, Darwin's book is titled 'On the Origin of the Species' Frank Sherwin, ICR PS I am in the process of writing a book for high school students tired of evolutionism. The book will answer some of the more outrageous macroevolutionary comments by Miller & Levine in their 2000 (5th ed) Biology textbook. Stay tuned. -----Original Message----- Sent: Monday, February 07, 2000 8:05 PM Subject: Yolk Sac in human embryos
|
|
|
|
|
The earliest events leading from the first division of the egg cell to the blastula stage in amphibians, reptiles and mammals |
The earliest events leading from the first division of the egg cell to the blastula stage in amphibians, reptiles and mammals are illustrated in Figure 5.4. Even to an untrained zoologist it is obvious that neither the blastula itself, nor the sequence of events which lead to its formation, is identical in any of the three vertebrate classes shown. The differences become even more striking in the next major phase in embryo formation - gastrulation. This process involves a complex sequence of relative cell movements whereby the cells of the blastula rearrange themselves, eventually resulting in the transformation of the blastula into the intricate folded form of the early embryo, or gastrula, which consists of the three basic germ cell layers: the ectoderm, which gives rise to the skin and the nervous system; the mesoderm, which gives rise to muscle and skeletal tissues; and the endoderm, which gives rise to the lining of the alimentary tract as well as to the liver and pancreas. No one doubts that gastrulation and the gastrula are homologous in all vertebrates, yet the way the gastrula is formed and particularly the positions in the blastula of the cells which give rise to the germ layers and their migration patterns during gastrulation differ markedly in the different vertebrate classes. In some ways the egg cell, blastula and gastrula stages in the different vertebrate classes are so dissimilar that, were it not for the close resemblance in the basic body plan of all adult vertebrates, it seems unlikely that they would have been classed as belonging to the same phylum. There is no question that, because of the great dissimilarity of the early stages of embryogenesis in the different vertebrate classes, organs and structures considered homologous in adult vertebrates cannot be traced back to homologous cells or regions in the earliest stages of embryogenesis. In other words, homologous structures are arrived at by different routes. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 145-6) |
|
|
|
|
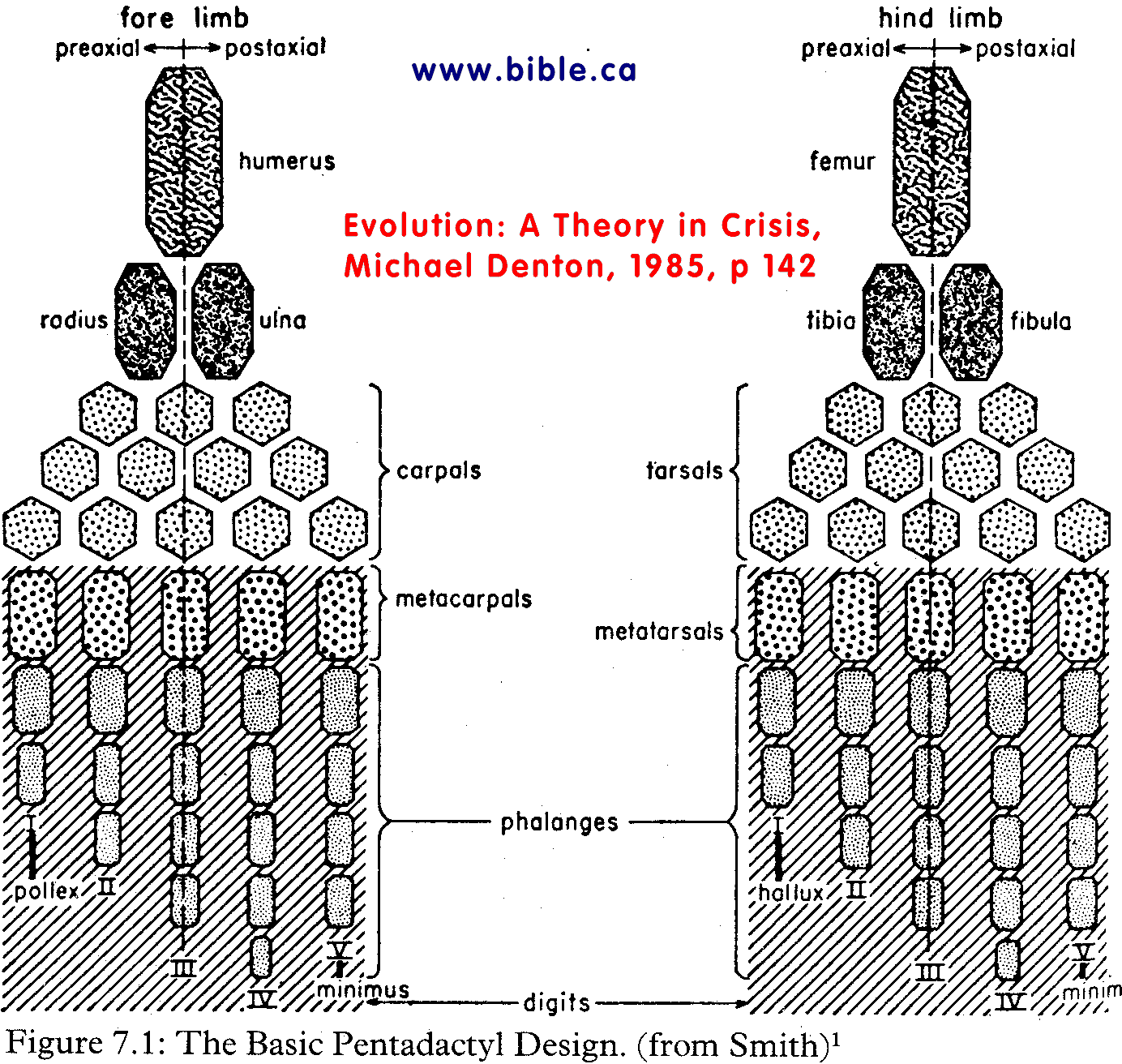
Pentadactyl design of vertebrate forelimbs and hindlimbs: |
"The evolutionary interpretation of homology is clouded even further by the uncomfortable fact that there are many cases of 'homologous like' resemblance which cannot by any stretch of the imagination be explained by descent from a common ancestor. The similar pentadactyl design of vertebrate fore- and hindlimbs provides the classic example. We have seen that the forelimbs of all terrestrial vertebrates are constructed according to the same pentadactyl design, and this is attributed by evolutionary biologists as showing that all have been derived from a common ancestral source. But the hindlimbs of all vertebrates also conform to the pentadactyl pattern and are strikingly similar to the forelimbs in bone structure and in their detailed embryological development. Yet no evolutionist claims that the hind- limb evolved from the forelimb, or that hindlimbs and forelimbs evolved from a common source. The striking similarity in the design of the fore- and hindlimbs of terrestrial vertebrates is seen in Figure 7.1. The detailed correspondence is remarkable. The proximal part of both the fore- and hindlimb is composed of one main bone, humerus in the arm, femur in the leg. The next section of the limbs is composed of two bones, radius and ulna in the arm, tibia and fibula in the leg. The hand and foot are also based on the same design, with five digits in both hand and foot. The first digit in hand and foot, the thumb and big toe, are both made up of only two small bones; the other digits are made up of three or more. There is no doubt that in terms of evolution the fore- and hindlimbs must have arisen independently, the former supposedly evolving from the pectoral fins of a fish, the latter from the pelvic fins. Here is a case of profound resemblance which cannot be explained in terms of a theory of descent. The occurrence of the same pentadactyl pattern in the fore- and hindlimbs presents an additional and unrelated challenge to evolutionary biology - that of explaining the independent origin of structures which are incredibly similar in terms of a random accumulation of tiny advantageous mutations. The adult form of the fore- and hindlimbs is not identical in any known vertebrate species. In every case the pentadactyl plan is considerably modified during development, so that the final adaptive form of both limbs is quite different and departs markedly from the basic pentadactyl plan, so much so that in many cases the original pentadactyl design is virtually impossible to detect in the final form of the limb. It seems very unlikely that there could be any adaptive necessity that dictates that there be five digits in both hand and foot or that thumb and big toe be both made up of two phalanges, that the forearm and lower leg be both made of two long bones or that there be only one bone in the upper arm and leg. We seem forced to propose that during the course of evolution the gradual accumulation of tiny independent and random changes in two independent structures - the pectoral and pelvic fins of a fish -hit on an identical yet apparently arbitrary ground plan for the design of the fore- and hindlimbs of a tetrapod. The problem is even more perplexing considering that neither the initial structures - the pelvic and pectoral fins of a fish - nor the end products of the process - the fore- and hindlimbs of a tetrapod - are in any strict sense identical. How this complex and seemingly arbitrary pattern was arrived at twice independently in the course of evolution is mystifying. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 153) |
|
|
|
|
vertebrate forelimbs |
Another class of organs considered strictly homologous are the vertebrate forelimbs yet they generally develop from different body segments in different vertebrate species. The forelimbs develop from the trunk segments 2, 3, 4 and 5 in the newt, segments 6, 7, 8 and 9 in the lizard and from segments 13, 14, 15, 16, 17 and 18 in man. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 146, citing: Homology: An Unsolved Problem, G. De Beer, 1971, p 8) |
|
|
|
|
vertebrate alimentary canal: |
Even after gastrulation the sites from which homologous structures are derived are different in different vertebrate classes. As De Beer points out, structures as obviously homologous as the vertebrate alimentary canal are formed from quite different embryological sites in different vertebrate classes. The alimentary canal is formed from the roof of the embryonic gut cavity in the sharks, from the floor in the lamprey, from roof and floor in frogs, and from the lower layer of the embryonic disc, the blastoderm, in birds and reptiles. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 146, citing: Homology: An Unsolved Problem, G. De Beer, 1971, p 13) |
|
|
|
|
occipital arch: |
Similarly, the position of the occipital arch relative to body segmentation varies widely in different vertebrate species. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 146, citing: Homology: An Unsolved Problem, G. De Beer, 1971) |
|
|
|
|
vertebrate kidney |
The development of the vertebrate kidney appears to provide another challenge to the assumption that homologous organs are generated from homologous embryonic tissues. In fish and amphibia the kidney is derived directly from an embryonic organ known as the mesonephros, while in reptiles and mammals the mesonephros degenerates towards the end of embryonic life and plays no role in the formation of the adult kidney, which is formed instead from a discrete spherical mass of mesodennal tissue, the metanephros, which develops quite independently from the mesonephros. Even the ureter, the duct which carries the urine from the kidney to the bladder, is formed in a completely different manner in reptiles and mammals from the equivalent duct in amphibia. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 146) |
|
|
|
|
The amniotic and allantoic membranes |
"A further example is provided by the development of the two unique membranes, the amniotic and allantoic, which surround the growing embryo in reptiles, birds and mammals. These membranes are considered to be strictly homologous in all the vertebrate groups in which they occur, but in mammals the processes which lead to their formation and the cells from which they are derived differ completely from those in reptiles and birds. In De Beer's words: "It does not seem to matter where in the egg or the embryo the living substance out of which homologous organs are formed comes from. Therefore, correspondence between homologous structures cannot be pressed back to similarity, of position of the cells of the embryo or the parts of the egg out of which these structures are ultimately differentiated. [emphasis added]" (Evolution: A Theory in Crisis, Michael Denton, 1985, p 147, citing: Homology: An Unsolved Problem, G. De Beer, 1971, p 13) |
|
|
|
|
Vertebrate eye |
In the same article De Beer goes on to describe in detail an interesting case which illustrates that even actual developmental mechanisms, by which apparently homologous structures are formed during embryogenesis, may not be homologous at all: "It was a problem to know why the lens of the vertebrate eye, which develops from the epidermis overlying the optic cup, should develop exactly in the "right" place, and fit into the optic cup so perfectly, until it was discovered that the optic cup is itself an organizer which induces the epidermis to differentiate into a tailor-made lens. At least, this is what it does in the common frog, Rana fusca, in the embyro of which, if the optic cup is cut out, no lens develops at all. But in the closely related edible frog, Rana esculents, the optic cup can be cut out from the embryo, and the lens develops all the same. It cannot be doubted that the lenses of these two species of frog are homologous, yet they differ completely in the mechanism by which determination and differentiation are brought about. This is no isolated example. In true vertebrates the spinal cord and brain develop as a result of induction by the underlying organizer; but in the "tadpole larva" of the tunicates, which has a "spinal cord" like the vertebrates, it differentiates without any underlying organizer at all. All this shows that homologous structures can owe their origin and stimulus to differentiate to different organizer- induction processes without forfeiting their homology. [emphasis added]" (Evolution: A Theory in Crisis, Michael Denton, 1985, p 147, citing: Homology: An Unsolved Problem, G. De Beer, 1971) |
|
|
|
|
Formation of insect organs through metamorphosis |
"Insect metamorphosis provides many other examples of homologous organs and structures being arrived at by radically different embryogenic routes. The first stage of metamorphosis, shortly following the formation of the pupa, involves what amounts to the virtual dissolution of all the organ systems of the larvae into a veritable soup of fragmented cells and tissues. This dissolution phase is quickly followed by an assembly phase during which all the organ systems - muscular, nervous and alimentary - of the adult insect are built up from special embryonic cells which occur either in specific places in the pupa, known as imaginal buds or discs, or scattered widely in the disintegrating tissues of the larva. (A General Textbook of Entomology, A. D. Imms, 1957, p 236) Detailed comparative studies of the processes of organ formation in different insect species have revealed that the ways in which the adult organ systems are formed during metamorphosis are bewilderingly diverse in different species. Take, for example, the formation of the alimentary tract. The lining of the midgut is always replaced during metamorphosis but the new adult midgut is reformed in some insects from primitive embryonic cells scattered throughout the old larval midgut while in other species by the migration of special embryonic cells from its posterior end. (A General Textbook of Entomology, A. D. Imms, 1957, p 240) The process of reconstruction of the fore- and hindguts also differs radically in different species. As entomologist Chapman describes it: (The Insects, R. F. Chapman, 1969, p 415) In Coleoptera (the beetles) the reconstruction of the stomatodoeurn (the foregut) and the proctodaeum (the hindgut) is carried out by the renewed activity of the larval cells without any accompanying cell destruction but in Lepidoptera (the butterflies) and Diptera (the flies) new structures develop from imaginal rings which are proliferating centres at the tips of the foregut and hindgut. Discussing the fate of the malphighian, or excretory, tubules of the larva, he continues: (The Insects, R. F. Chapman, 1969, p 416) In Coleoptera the tubules are rebuilt from special cells in the larval tubules while in Hymenoptera (the ants and bees) the larval tubules break down completely and are replaced by new ones developing from the tip of the proctodaeum (the hindgut). No one would doubt that the alimentary tract and the malphighian tubules are homologous in all insect species but, again as in the vertebrate cases cited above, we see that "homology" cannot be traced back to similar embryogenic processes and events. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 147-8) |
|
|
|
|
homology in plants |
The same principle also holds in the case of many sorts of homology in plants. For example, the seeds of the conifers and the flowering plants (the angiosperms) are considered to be homologous by most botanists, and indeed the close resemblance in the structure of the seeds in both groups is used by taxonomists as one of the key character traits to classify them together in the major group Spermatophyta. Each seed consists of an enclosed egg cell or ovule plus a food store (the endosperm) which surrounds the ovule and supplies nourishment to the growing embryo after fertilization. Yet the way in which the ovule and endosperm are formed profoundly differs in the two groups in a number of important respects. (Evolution: A Theory in Crisis, Michael Denton, 1985, p 148, citing: The Living Plant, A. J. Brook, 1964, p 281,281,466) |
|
|
|
|
apparently homologous structures are specified by quite different genes in different species |
"The evolutionary basis of homology is perhaps even more severely damaged by the discovery that apparently homologous structures are specified by quite different genes in different species. The effects of genes on development are often surprisingly diverse. In the house mouse, nearly every coat-colour gene has some effect on body size. Out of seventeen x-ray induced eye colour mutations in the fruit fly Drosophda melanogaster, fourteen affected the shape of the sex organs of the female, a characteristic that one would have thought was quite unrelated to eye colour. Almost every gene that has been studied in higher organisms has been found to effect more than one organ system, a multiple effect which is known as pleiotropy. As Mayr argues in Population, Species and Evolution. "It is doubtful whether any genes that are not pleiotropic exist in higher organisms. Since the primary gene action in multicellular organisms is usually several steps removed from the peripheral phenotypic character, it is obvious that non pleiotropic genes must be rare if they exist at all." (Populations, Species and Evolution, E. Mayr, 1970, p 93) Not only are most genes in higher organisms plieotropic in their influence on development but, as is clear from a wide variety of studies of mutational patterns in different species, the pheotropic effects are invariably species specific. In Figure 7.2 the multiple effects of one particular gene in the domestic chicken arc illustrated. As can be seen, a mutation in this gene causes developmental abnormalities in a variety of systems. Here then is a gene that is involved in the development of some structures unique to birds - air sacs and downy feathers - and of other structures such as lungs and kidneys, which occur in many other vertebrate classes. This can only mean that non-homologous genes are involved to some extent in the specification of homologous structures. Another simple example is a gene, again in the domestic fowl, which controls the formation of the crest of feathers and causes a cerebral hernia with upswelling of the skull in the form of a knob to accommodate it. It is difficult to believe that this gene has any homologue in vertebrate species lacking feathers, and yet it is involved in the development of the skull, a feature possessed by all vertebrate species. As De Beer says, "Homologous structures need not be controlled by identical genes and homology of phenotypes does not imply similarity of genotype." (Evolution: A Theory in Crisis, Michael Denton, 1985, p 150, citing: Homology: An Unsolved Problem, G. De Beer, 1971, p 15) |
|
|
|
|
Many other examples of this sort of the phenomenon |
There are many other examples of this sort of the phenomenon, adaptations of great complexity which exhibit very close resemblance in their design but which must have arisen entirely independently. Such examples of convergence led Carter to comment: "There are many problems in evolution for which our present explanations are inadequate or incomplete. This is certainly one place in which this is so. It is clear that much more work must be done before we have a complete understanding of the process of evolution." (Evolution: A Theory in Crisis, Michael Denton, 1985, p 153, citing: Structure and Habit in Vertebrate Evolution, G. S. Carter, 1967, p 493) |
|
|
|
|
Photo Gallery of the Homology Fraud |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Evolutionists arguments and rebuttals |
Evolutionist argument:
"If you look at a 1953 Corvette and compare it to the latest model, only the most general resemblances are evident, but if you compare a 1953 and a 1954 Corvette, side by side, then a 1954 and a 1955 model, and so on, the descent with modification is overwhelmingly obvious. This is what paleontologists do with fossils, and the evidence is so solid and comprehensive that it cannot be denied by reasonable people.." (Evolution and the Myth of Creationism, Tim Berra, 1990, p 117)